















 25 / 32
25 / 32

4.1
Trasmissione sinaptica
69
antagonisti, viene favorito lo stato inattivo del recettore
che impedisce l’attivazione delle proteine G. In seguito
all’attivazione della proteina G, una molecola di GTP
spiazza il GDP dal suo sito di legame; contemporane-
amente la subunità
a
si stacca dal complesso
bg
della
proteina G. La subunità
a
e il complesso
bg
possono
interagire con canali ionici (
Fig. 4.7A
) o enzimi intra-
cellulari (
Fig. 4.7B
).
RIMOZIONE DEL
NEUROTRASMETTITORE DALLA
FESSURA SINAPTICA
L’interruzione della trasmissione sinaptica inizia
quando termina l’esocitosi del neurotrasmettitore ma
per completarsi essa richiede anche l’eliminazione del
neurotrasmettitore ancora presente nella fessura sinap-
tica. La rimozione del mediatore chimico può avvenire
attraverso tre meccanismi che, secondo la sinapsi con-
siderata, hanno diversa importanza. Essi sono:
1) la
diusione del neurotrasmettitore
fuori dalla fes-
sura sinaptica per gradiente di concentrazione;
2)
l’inattivazione enzimatica del neurotrasmettitore
ad
opera di enzimi idrolitici localizzati nella fessura
sinaptica (membrana postsinaptica);
3) il
recupero (“re-uptake”) del neurotrasmettitore
da
parte dell’elemento presinaptico grazie alla pre-
senza di meccanismi di trasporto dedicati che per-
mettono sia la rimozione del neurotrasmettitore
dalla fessura sinaptica sia il suo riutilizzo da parte
dell’elemento presinaptico.
SINAPSI ECCITATORIE E SINAPSI
INIBITORIE
Sia nelle sinapsi chimiche veloci che nelle sinapsi chi-
miche lente, le caratteristiche elettriche del potenziale
postsinaptico dipendono dalle proprietà bio"siche dei
canali ionici attivati direttamente o indirettamente
dal neurotrasmettitore. I canali ionici attraverso i
quali operano i neurotrasmettitori sono canali catio-
nici (recettori canali) con una permeabilità prevalente
per il Na
+
, K
+
o Ca
2+
oppure canali anionici per il Cl
–
. In
seguito alla loro apertura, la variazione del potenziale
di membrana della cellula postsinaptica sarà dettata dai
potenziali d’equilibrio degli ioni che permeano attra-
verso i canali e dal potenziale di membrana a riposo
(
V
r
) della cellula postsinaptica. Ad esempio, assumendo
i potenziali di equilibrio pari a
E
Na
= +62 mV,
E
K
= –102
mV,
E
Ca
= +125 mV ed
E
Cl
= – 88 mV ed il valore di
V
r
della cellula postsinaptica pari a – 80 mV, i gradienti
elettrochimici per il Na
+
, Ca
2+
e Cl
–
guideranno gli ioni
a di#ondere verso l’interno della cellulamentre il $usso
di ioni K
+
sarà diretto verso l’esterno (Cap. 3). Di con-
seguenza, se il neurotrasmettitore attiva direttamente
o indirettamente un canale ionico con permeabilità
prevalente per il Na
+
o per il Ca
2+
, i canali attivati rende-
ranno più positivo l’interno della cellula postsinaptica
(per ingresso di cationi) e la variazione transiente del
potenziale dellamembrana postsinaptica sarà in senso
depolarizzante
(
Fig. 4.8A
). Al contrario, se il neurotra-
smettitore controlla l’apertura di un canale ionico con
permeabilità prevalente per il K
+
o selettivo per il Cl
–
,
il potenziale di membrana della cellula postsinaptica
cambierà in senso
iperpolarizzante
,
rispettivamente
a causa dell’uscita di cationi o dell’ingresso di anioni
(
Fig. 4.8B
).
Quando il neurotrasmettitore causa depolarizza-
zione della cellula postsinaptica, il neurotrasmettitore,
il potenziale postsinaptico e la sinapsi chimica sono
de"niti
eccitatori
. In tal caso, il neurotrasmettitore
causa un avvicinamento del potenziale di membrana
postsinaptico al valore di soglia. Se il neurotrasmetti-
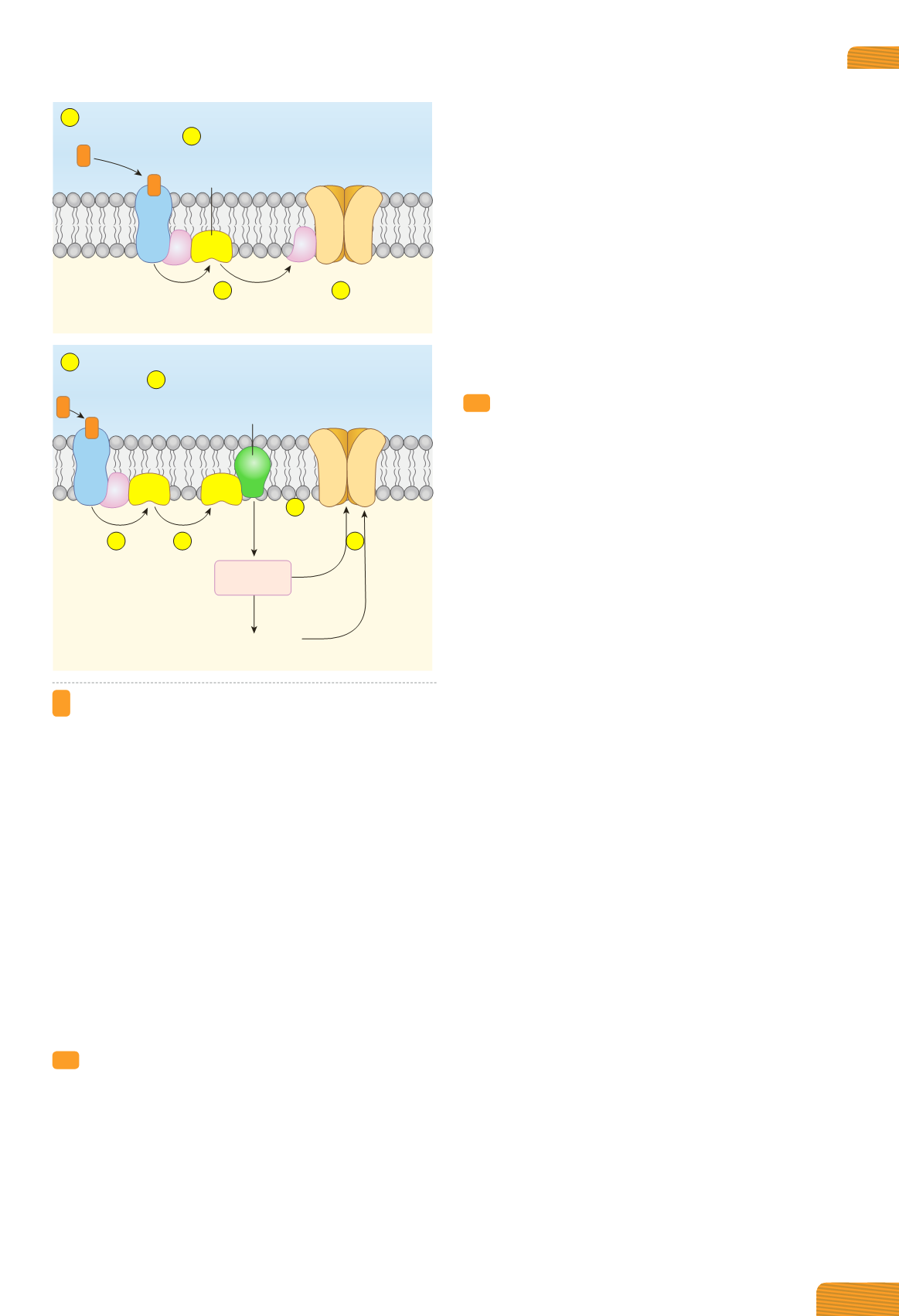
Figura 4.7
Recettori metabotropi.
(A)
La proteina G attivata
può modulare direttamente l’attività di un canale ionico. Il
processo è limitato alla membrana cellulare e vede implicati:
il recettore, la proteina G attivata e il canale ionico (e=et-
tore).
(B)
La proteina G attivata può controllare l’attività di
un enzima intracellulare. In tal caso, il processo è più com-
plesso ed è mediato da secondi messaggeri, che esercitano
la loro azione su canali ionici direttamente o mediante il
coinvolgimento di protein chinasi.
Il neurotrasmettitore
si lega al recettore
Risposte lente, accoppiamento diretto
Risposte lente, intervento di secondi messaggeri
Proteina G
`
a
`
a
_
R
Apertura o chiusura
del canale ionico
Attivazione
della proteina G
1
1
2
3
Il neurotrasmettitore
(primo messaggero)
lega il recettore Enzima
` _
_
R
Apertura
o chiusura
di canali
ionici
Attiva
la proteina
G
2
Attiva
o inibisce
l'enzima
3
Produce
secondo
messaggero
4
5
Protein chinasi
Secondo
messaggero
a
A
B