















 31 / 32
31 / 32

4.1
Trasmissione sinaptica
75
riali per l’ACh, i cui membri dieriscono per struttura,
proprietà farmacologiche e distribuzione tissutale. Per
tale motivo, per denirlo inequivocabilmente, l’esatta
denominazione del canale acetilcolinico presente nella
giunzione neuromuscolare è
recettore-canale nicoti-
nico di tipomuscolare
, universalmente abbreviato con
nAChR
(
Nicotinic Acetyl-Choline-Receptor
) muscolare.
Come ipotizzato da Fatt e Katz, il nAChR musco-
lare è localizzato prevalentemente nella regione di
placca. La distribuzione dei nAChR nella giunzione
neuromuscolare è stata ampiamente documentata
con tecniche istochimiche o autoradiograche e l’im-
piego della
a
-bungarotossina, una tossina contenuta nel
veleno di alcuni cobra acquatici (ad esempio
Bungarus
multicinctus
), che si lega selettivamente e irreversibil-
mente ai nAChR. In particolare, il nAChR è presente
sulle creste delle pieghe giunzionali dove raggiunge
densità di 10.000-15.000 unità/
m
m
2
(contro le 5 unità/
m
m
2
delle regioni extragiunzionali). Sul fondo delle pie-
ghe si trovano invece localizzati i canali del Na
+
e del
K
+
voltaggio-dipendenti che convertono il potenziale
postsinaptico in un potenziale d’azione.
Il legame di due molecole di ACh al nAChRmusco-
lare causa l’apertura del recettore-canale. Quando il
nAChRmuscolare è aperto, il poro ionico è attraversato
da una corrente cationica entrante (
Fig. 4.14A
). Essa è
prevalentemente generata da ioni Na + (!usso entrante)
e K
+
(!usso uscente). Altri cationi che permeano il
canale sono gli ioni Ca
2+
che entrano all’interno della
cellula. Sebbene il Ca
2+
possa svolgere importanti
funzioni di modulazione sul canale stesso (sia in ter-
mini di proprietà biosiche sia in termini di controllo
di espressione), la permeabilità al Ca
2+
è così ridotta
che il contributo di questo ione nella generazione del
potenziale postsinaptico è trascurabile.
Si è già detto che il potenziale d’azione generato
dal motoneurone causa la liberazione di una quantità
elevatissima di ACh nella fessura sinaptica (corrispon-
dente a circa 100-200 quanti). È stato stimato che il neu-
rotrasmettitore nella fessura sinaptica raggiunge una
concentrazione dell’ordine delle millimoli. L’elevata
concentrazione del neurotrasmettitore e l’elevata den-
sità di recettori sulle creste giunzionali garantiscono
l’apertura contemporanea di moltissimi nAChR e la
generazione di un potenziale postsinaptico eccitatorio
abbondantemente sovrasoglia. Quindi, ad ogni poten-
ziale d’azione neuronale (presinaptico) corrisponde un
potenziale d’azione muscolare (postsinaptico) capace
di propagarsi lungo la bra muscolare scheletrica e di
causarne la contrazione (
Fig. 4.14B
).
L’elevata e$cienza della trasmissione sinaptica è
una peculiarità della giunzione neuromuscolare; essa è
assicurata dall’ampia supercie di contatto delle mem-
brane sinaptiche che da un lato rendono possibile la
liberazione di numerose vescicole sinaptiche e dall’al-
tro assicurano una straordinaria ricettività chimica da
Numero di eventi
Numero di eventi
Insuccessi
18
16
14
12
10
8
6
4
2
40
30
20
10
Ampiezza dei potenziali di placca in miniatura (mV)
0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0
0,2 0,4
Ampiezza dei potenziali di placca
in miniatura (mV)
0,6 0,8
2,2 2,4 2,6 2,8 3,0
0 0,2 0,4
I
II
III
IV
V
VI
VII
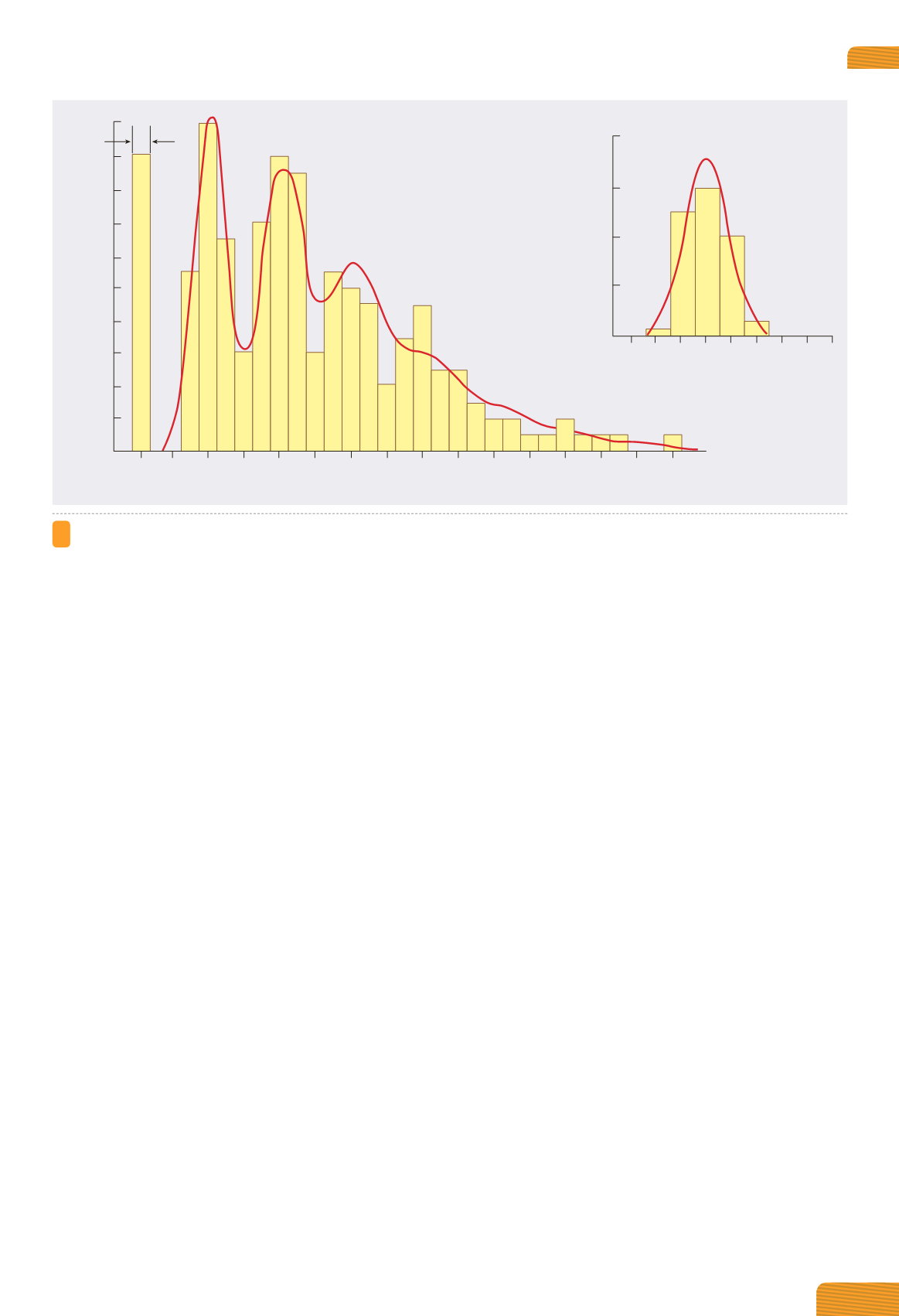
Figura 4.13
Distribuzione delle ampiezze dei potenziali postsinaptici. L’istogramma mostra le ampiezze dei potenziali postsinaptici
misurati in una sinapsi neuromuscolare. Si noti che i picchi delle ampiezze (0,4, 0,8, 1,2, …) corrispondono a multipli interi dell’am-
piezza media dei potenziali in miniatura (inserto). La curva rappresenta l’andamento teorico della distribuzione delle ampiezze,
calcolato mediante l’equazione di Poisson, assumendo che i potenziali postsinaptici corrispondono alla somma di più miniature
indipendenti.