















 24 / 32
24 / 32

TRASMISSIONE SINAPTICA
68
CAPITOLO 4
causa diretta della variazione del potenziale di mem-
brana della cellula postsinaptica e la comunicazione
è denita
trasmissione diretta
o
sinapsi chimica veloce
(
Fig. 4.6
). Analizzando le sequenze aminoacidiche dei
recettori ionotropi clonati è emersa una struttura simile
comune, anche se probabilmente due diversi geni ance-
strali hanno dato origine a due famiglie recettoriali. Alla
prima famiglia appartengono il recettore nicotinico per
l’acetilcolina, il recettore per l’acido
a
- aminobutirrico
GABA
A
, il recettore per la glicina, alcuni recettori per
la serotonina. Alla seconda, i recettori ionotropi per il
glutammato. Da un punto di vista strutturale, i recettori
ionotropi sono costituiti da quattro o cinque subunità
proteiche, ciascuna delle quali contribuisce a formare
il poro del canale.
In altre sinapsi, il neurotrasmettitore si lega a un
recettore accoppiato a una proteina G (
recettoremeta-
botropo
o
metabotropico
) che a sua volta modula l’at-
tività di un canale ionico o direttamente o attraverso
l’attivazione di un secondo messaggero. In entrambi i
casi, la variazione della permeabilità della membrana
postsinaptica richiede il susseguirsi in cascata di diversi
eventi citoplasmatici e si parla di
trasmissione indi-
retta
o di
sinapsi chimica lenta
. I recettori metabotropi
sono una classe molto più numerosa e sono proteine
transmembrana a 7
a
-eliche che non formano canali
ionici. Hanno un sito di legame extracellulare per il
neurotrasmettitore e una regione intracellulare depu-
tata al legame con le proteine G, le quali sono costituite
da tre subunità (
a
,
b
,
g
) che legano nucleotidi guani-
nici. Quando una proteina G viene in contatto con un
recettore metabotropo occupato dal suo agonista (stato
attivo), interagisce con il complesso recettore-agonista
e viene attivata. Nel caso di un recettore occupato da
Ca
2+
Ca
2+
A
B
C
D
E
mV
mV
mV
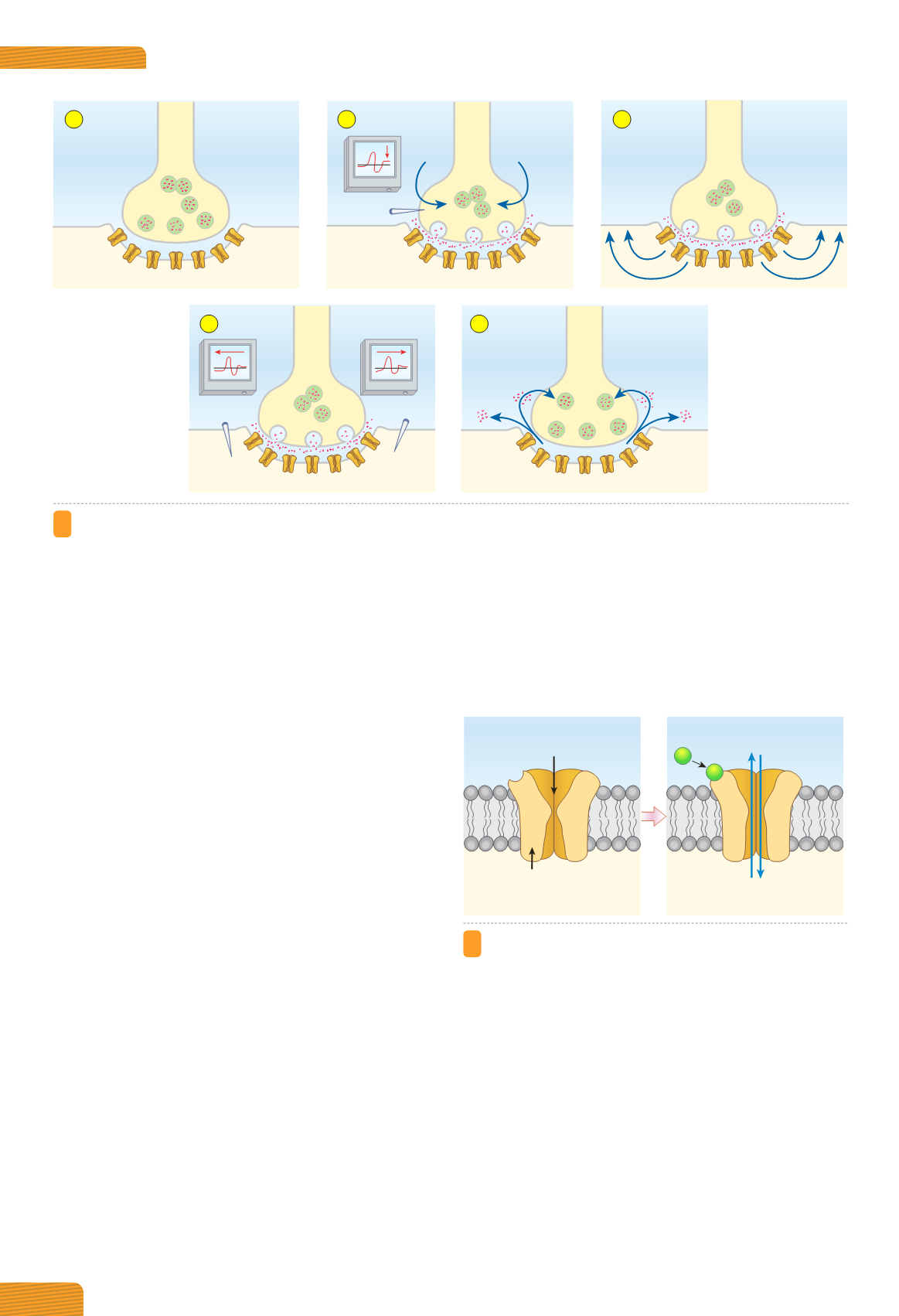
Figura 4.5
Fasi della trasmissione chimica.
(A)
Nella sinapsi chimica, a riposo, il neurotrasmettitore si trova racchiuso nelle
vescicole sinaptiche.
(B)
La trasmissione sinaptica chimica inizia con la liberazione del neurotrasmettitore indotta dal potenziale
d’azione presinaptico. Questo processo è Ca
2+
-dipendente e richiede l’apertura dei canali del Ca
2+
voltaggio-dipendenti presenti
nella membrana della terminazione presinaptica.
(C)
Il neurotrasmettitore diAonde attraverso la fessura sinaptica e si lega a spe-
cifiche molecole recettoriali presenti nella membrana della cellula postsinaptica. Il legame del neurotrasmettitore ai rispettivi
recettori causa una variazione del potenziale circoscritto alla regione della membrana sinaptica in cui i recettori sono presenti.
Successivamente, si generano correnti elettrotoniche tra la membrana sinaptica e le regioni di membrana elettricamente eccitabili
limitrofe, dove sono presenti i canali del Na
+
e del K
+
voltaggio-dipendenti.
(D)
Se l’intensità delle correnti elettrotoniche rende
il potenziale della membrana eccitabile pari o superiore al valore soglia, nella cellula postsinaptica insorge il potenziale d’azione.
(E)
La trasmissione termina con l’eliminazione del neurotrasmettitore dalla fessura sinaptica.
Figura 4.6
Recettori ionotropi. Il legame del neurotrasmet-
titore nel sito specifico del canale ligando-dipendente deter-
mina una modificazione strutturale della proteina generando
un flusso ionico che altera in modo transiente le proprietà
elettriche della cellula postsinaptica.
Fluido extracellulare
Legame
del neurotrasmettitore
Flusso ionico
Canale ionico
Recettore ionotropo
Citosol